Joel Brind, Ph.D
http://180degreehealth.com/diet-inflammation-part/
Diet and Inflammation Part 1
Jul 24, 2014
There’s no shortage of info out there about inflammation’s being the root cause of most of what makes people sick and die these days–from arthritis to diabetes to cardiovascular disease to cancer—and what to eat and not eat to put your body into an “anti-inflammatory†state.
But there is a shortage of real knowledge about inflammation itself: What its proper role is in the body and why it is so often out of control. Once you know that, the rest is easy.
Back in the day (some 35 years ago), I studied immunology at NYU Medical School as part of my graduate training in Basic Medical Science. You would think that inflammation, as a major function of the immune system—innate immunity, to be precise—would occupy a substantial proportion of the course material. Nope! Everyone in the field, it seemed, wanted to win the Nobel Prize for figuring out acquired immunity: How does the body figure out how to make a specific antibody against the chemical signature of a microbe it has never encountered before? That was one of the great mysteries of the day; the principle behind vaccination and all manner of specific immunity. Well, that mystery was eventually solved, and I don’t even remember who got the credit. Are we making better vaccines and saving more lives because of it? You be the judge.
But if you go back to the days when infectious disease killed most people; when great plagues swept through human populations with some regularity, it wasn’t acquired immunity that saved most of the survivors. It was innate immunity. After all, acquired immunity takes weeks to develop after the first exposure, and the body can succumb to infection in a few days. So the real life saver is the first response system, the aggressive, non-specific attack against the offending microbes. Of course, such attack can and does do lots of damage to normal tissues in the process, but it might just save your life.
It’s really just like society’s first responders; like the fire department. When your fire alarm detects a fire, the fire department is summoned, and quickly the firemen show up with their axes and their hoses. They put the fire out, all right, but they do lots of damage in the process. Once the fire is out, you bring in the repair contractor to fix the damage. In your body, once the threat of infection has been neutralized, healing begins.
But for some odd reason, inflammation—the basic operation of innate immunity—is still viewed by the medical and scientific community as part of the healing response. In fact, the opposite is true: it inhibits healing quite effectively.
Two years ago, this is how Wendy Weber, A National Center for Complementary and Alternative Medicine (one of the NIH Institutes) program director, was quoted in the Wall Street Journal (wsj.com; the section entitled “The Informed Patientâ€, article author Laura Landro). ‘â€You need to have inflammation when you have a wound and the immune system goes in to heal it. Yet we don’t want too much inflammation in our system causing damage to our arteries†and other harm’.
The prevailing dogma, unfortunately, does not really distinguish the separate roles of immune defense and tissue repair.
To take a simple example, say you sprain your ankle. It gets all swollen and painful and immobilized from what process? Inflammation. In fact, everyone knows that the immediate treatment of choice is to put ice on it. To do what? To suppress inflammation!
Why? Because the inflammation prevents healing from taking place.
OK, so why does inflammation happen at all when you sprain your ankle? Because, it is generally believed, inflammation is the body’s natural response to tissue injury; to distress signals released from damaged cells and tissues. But why? If you sprain your ankle, where is the route of infection? There are no invading microbes to destroy, and if inflammation happens anyway and does more harm than good, why does your body do it? Why do you have to interfere with your body’s natural response to the blunt injury, for optimal healing to occur?
The answer is quite simple: Your body is not acting appropriately, because it is laboring under a nutritional imbalance. Actually, the typical Western diet tends to be deficient and/or imbalancedin 3 key nutrients: salicylic acid, omega-3 fatty acids (vs. omega 6), and glycine (vs. methionine). The third is the most important and the least well known (although Vladimir Heiskanen’s excellent recent post on this blog gives a pretty good introduction). I’ll get to the specifics of how all this works—down to the cellular and molecular level–in my next post.
Diet and Inflammation Part 2
http://180degreehealth.com/diet-inflammation-part-2/
Aug 11, 2014
My last post—part 1 of Diet and Inflammation—left off with my conclusion that the answer to the widespread prevalence of chronic inflammation was a nutritional imbalance incurred by the typical Western diet, specifically deficiencies or imbalances in 3 key nutrients: omega-3 (v. omega 6) fatty acids, salicylic acid and glycine (v. methionine).
What exactly happens in inflammation? Inflammation is the basic action of the innate immune system to destroy potentially pathogenic microbes that get into the internal body tissues. It is a non-specific aggressive action by amoeba-like cells called macrophages (derived from the Greek and meaning “big eatersâ€), the immune system’s first responders, which can literally gobble up bacteria and other microbes. Several types of these cells—called granulocytes—have long been recognized as circulating in the blood as a type of white blood cell. More recently, macrophages have been recognized as populating all sorts of organs and tissues.
But all these diverse types of macrophages are immune system cells, and all of them originate in the bone marrow. If there is tissue injury, injured cells and cell debris will be gobbled up by these macrophages. But if there is infection—the recognition of the generic signature of bacteria, for example—these macrophages get activated, producing toxins such as hydrogen peroxide in order to kill the bacteria. In order to amplify the reaction, these macrophages release chemical signals called prostaglandins to recruit and activate other macrophages, the purpose being to destroy the infecting microbes before they can destroy the host. Of course, like other first responders, the macrophages will put down the infection just like the firemen will put out the fire, but they will also do lots of damage to normal tissues.
Unfortunately, there is still lots of confusion out there about the role of inflammation and innate immunity; most medical authorities believing that inflammation is part of the healing response and a normal response to tissue injury. Why? Because it always seems to happen with tissue injury. You sprain your ankle, and it gets swollen and painful and immobilized; i.e., inflamed. But why should this happen if there is no route of infection? There are no microbes to kill. And we all know that, contrary to healing, the inflammation inhibits healing, which is why we have to put ice on the injury, to suppress inflammation. So why does your body do it?
Here’s where nutrition comes in. The activation of macrophages is affected by an electrochemical switch mechanism on the cell surface membrane. When these cells are at rest, they are, so to speak, switched off, there is a resting voltage between the outside and the inside of the cell (positive outside; negative inside). Just like a light switch on your wall, when it is off, there is a resting voltage (120 volts in the US) between the hot wire attached to the switch and the light fixture. When the light is switched on, the voltage drops as the energetic electrons flow through the switch and activate the fixture. The electrochemical switches in cells are channels in the membrane which allow positively charged ions (calcium or sodium ions) to flow across the membrane. The voltage drops when these channels open up momentarily, activating the cell. That’s how nerve impulses (called action potentials) activate muscles, for example. But it’s also how macrophages get activated.
However, the cell surface membrane is a very dynamic envelope, like a constantly moving, constantly changing, highly sophisticated soap bubble. In the course of ordinary activity, lots of leakage of ions occurs, and there are specialized channels in the membrane that let negatively charged chloride ions in to maintain the resting voltage of 0.07 volts. A substantial proportion of these chloride channels are operated—i.e., maintained in an open position—by the amino acid glycine. Glycine is ordinarily present in body fluids at high concentrations. But if they are not high enough, the glycine-gated chloride channels (aka glycine receptors) are not open enough to allow adequate chloride entry. Thus, the voltage between the outside and inside of the cell deteriorates and the cell is too easily activated, like when there is tissue injury but no infection. Once a macrophage is activated, the extent to which it recruits and activates other macrophages to the site of inflammation is related to the levels of the two other key nutrients: salicylic acid and omega-3 v. omega-6 fatty acids.
How the balance—or imbalance—in the intake of these types of polyunsaturated fatty acids (PUFAs) will be the subject of Part 3 in this series.
Comments:
Brad on August 11, 2014 at 3:57 pm
There’s a lot of buzz about gelatin these days either from bone broth or gelatin/collagen supplements. Do you believe an individual should supplement with a bone broth/gelatin/collagen that provides plenty of glycine especially in ratio to Methionine, or is it more advantageous to supplement with pure Glycine? I was curious if glycine works synergistically with other amino acids and if it’s important to have them properly balanced or if supplementing solely with pure Glycine will provide the anti-inflammatory benefits. Thanks.
Matt Stone on August 12, 2014 at 3:19 pm
Ray Peat states this about this very topic Brad…
A generous supply of glycine/gelatin, against a balanced background of amino acids, has a great variety of anti-stress actions. Glycine is recognized as an “inhibitory†neurotransmitter, and promotes natural sleep. Used as a supplement, it has helped to promote recovery from strokes and seizures, and to improve learning and memory. But in every type of cell, it apparently has the same kind of quieting, protective anti-stress action. The range of injuries produced by an excess of tryptophan and serotonin seems to be prevented or corrected by a generous supply of glycine. Fibrosis, free radical damage, inflammation, cell death from ATP depletion or calcium overload, mitochondrial damage, diabetes, etc., can be prevented or alleviated by glycine.
Some types of cell damage are prevented almost as well by alanine and proline as by glycine, so the use of gelatin, rather than glycine, is preferable, especially when the gelatin is associated with its normal biochemicals. For example, skin is a rich source of steroid hormones, and cartilage contains “Mead acid,†which is itself antiinflammatory.
====================================================
http://180degreehealth.com/diet-inflammation-part-3/
Diet and Inflammation Part 3
Aug 25, 2014
In my last post—Part 2 of this diet and inflammation series—I discussed the cells—called macrophages—which actually affect the inflammatory response, and how the amino acid glycine is crucial in regulating the activation of the macrophages at the level of the cell surface membrane. In the present installment, I’ll be discussing the propagation and amplification of the inflammatory response, and the key roles played by two other nutrients: salicylic acid and omega-3 (v. omega-6) fatty acids.
The cell membrane itself is made up mainly of molecules called phospholipids; natural soap-like molecules which each contain two fatty acids. Upon cellular activation, some of these phospholipid molecules are broken down such that one of the fatty acids is enzymatically converted to a messenger molecule—a prostaglandin—which diffuses away to activate—or inhibit the activation of—other cells it reaches (Such local messenger molecules are known as paracrine factors.) The enzyme that catalyzes the key step in the process is called a cyclooxygenase 2 (COX2). Salicylic acid inhibits the activity of COX2. (The synthetic drug aspirin, or acetylsalicylic acid, is a much more potent COX2 inhibitor; more on this later.)
The type of prostaglandin molecule released reflects the composition of the type of fatty acids that make up the membrane, which in turn reflects the fatty acid composition of the diet. The prostaglandins that amplify the inflammatory activation are made from the omega-6 fatty acid, arachidonic acid (AA); whereas prostaglandins that inhibit this activation are made from the omega-3 fatty acid, eicosapentaenoic acid (EPA).
Hence, the greater the preponderance of omega-6 fatty acids in the diet (largely from seed oils, e.g., corn, soybean, sunflower, peanut) as opposed to omega-3 (fish or krill oil, flaxseed oil), the greater the amplification of the pro-inflammatory prostaglandin signal. (The optimal dietary ratio of omega-6 to omega-3 is about 3 or 4 to one; although whether either type of these polyunsaturated fatty acid types—â€PUFAsâ€â€”are even essential to the human diet is still debated. Oils that seem to have a perfect balance of fatty acid types include walnut and olive oils [Hemp Seed Oil ~JP].) Since the mass campaign to replace saturated fats with PUFAs over the latter half of the 20th century was largely successful in saturating the Western diet (and that of its livestock) with omega-6 PUFAs, it has clearly contributed to the high prevalence of chronic inflammation.
Meanwhile, the extent of COX2 activity is largely controlled by the concentration of salicylic acid. For some reason, salicylic acid is often viewed as a “nutraceuticalâ€, rather than an everyday nutrient. In fact, it is often not viewed as a nutrient at all, although aspirin is viewed as something middle-aged and older individual are encouraged to take daily to prevent thrombotic events such as heart attacks and strokes. Aspirin, however, is a potent synthetic drug. Although it acts like salicylic acid (as well as increasing the actual salicylic acid content of the blood), it has potentially dangerous side-effects, like excess bleeding. Meanwhile, salicylic acid itself is a widespread botanical compound, particularly high in berry fruits, grapes and kiwis, and also present in significant amounts in nuts like almonds and walnuts. (It is also a key component of EVOO that is removed when olive oil is refined.)
It is my belief that it is salicylic acid, rather than the much touted polyphenols in fruits and nuts which are key to “anti-inflammatory dietsâ€, and which lower risk of cardiovascular disease, for example. These polyphenols (e.g., resveratrol, quercetin) are great anti-oxidants, but the value of anti-oxidants is largely to mitigate the effects of inflammation. I think it’s better to stop inappropriate inflammation from getting started. I also believe, for example, that the “French paradoxâ€â€”why French people eat such a high-fat diet but suffer a low rate of heart disease—is not due to the polyphenols in the wine they drink daily, but from the high salicylic acid content of grapes, and therefore, wine.
So it’s pretty clear how the typical Western diet that is low in fruits and vegetables, and low in omega-3—but high in omega-6 fats—contributes to excess inflammation, by helping to amplify—and therefore exaggerate—the inflammatory response. But getting back to the initiation of inflammation in the first place, why should glycine levels be low in the first place, and allow inflammation to develop inappropriately? After all, glycine is a non-essential amino acid, so you really should not have to eat any of it, right? And if the diet is rich in high-protein foods (meat, fish, poultry, eggs, dairy), we are also eating plenty of it.
The answer turns out to be quite simple: Although we discovered a century ago that we need to eat whole grains to avoid devastating deficiency diseases like pellagra, we never thought that we also need to eat whole cows, pigs, chickens and fish! But it turns out that the key to a healthy omnivorous diet is to balance the content of essential amino acid methionine that predominates in muscle with the glycine that predominates in the bones and connective tissues, the parts we usually throw away.
The specifics of the biochemistry—the metabolic interactions of glycine and methionine and key intermediate metabolites and cofactors—are now understood, and present a fascinating picture of how our bodies’ metabolic machinery works as best it can with what we feed it to keep us alive and healthy. That will be the focus of my next post.
Comments:
As for sinus infections, I have been troubled by that at times, during the winter months. I assumed it was cold sensitivity, and glycine helps in that it keeps the fluids thin, as there is lots of fluid but no inflammation. Inflammation is what results in the really thick mucus that generates secondary infections.
http://180degreehealth.com/diet-inflammation-part-4/
Diet and Inflammation Part 4
Sep 20, 2014
My last post focused on the propagation of the inflammatory signal, and how nutrients such as omega-6 PUFAs and the lack of salicylic acid results in amplification of that signal, thus contributing to the overall increase in inflammation-related disease we currently experience.

Collagen
But what about the initiation of the inflammatory signal in the first place? That often turns out to be due to a widespread deficiency in the amino acid glycine, which I described in an earlier post. Glycine acts as a sort of “cellular voltage regulatorâ€, preventing the inappropriate initiation of inflammation in response to cellular injury. (Inflammation is really only appropriate when there is infection taking place.)
But if glycine is the most abundant amino acid in the body, and it is nonessential—i.e., the human body can make it from scratch—why should anyone be deficient, especially nowadays, when the dietary intake of protein is usually so high?
As Matt Stone often says, it is really a matter of context; a matter of balance, rather than a matter of absolutes. Glycine is so abundant that it comprises about 22% by weight of the most abundant protein in the body. That protein would be collagen, the tough, extracellular fibrous protein that makes up the bones, cartilage and all the connective tissues. Hence, the collagen—and therefore most of the glycine—is the part of the meat, fish and poultry that we usually throw away. Bone broth is one way to recover it in the diet, and when the collagen is boiled out of the bones and purified, it is called gelatin.
But even though we discard most of the glycine from our animal flesh foods, we are still taking some in with our muscle meats, so that still doesn’t explain a widespread glycine deficiency. But in fact, the consumption of muscle meats actually exacerbates the deficiency because of the amino acid content of the muscle meats. Specifically, muscle meats are very rich in the essential amino acid methionine, and it is an understanding of the intimate relationship between methionine and glycine that provides the answer to this question of balance.
A problem with traditional nutritional thinking on amino acids is the rigid classification of those which are essential, as opposed to those which are non-essential; in particular, a disproportionate interest in the former v. the latter. Methionine, in particular, has long been known to play key roles in metabolism independent of its role as a constituent of proteins. Specifically, methionine—when activated to form S-adenosylmethionine (SAMe), is the universal methyl group donor, which adds a one-carbon methyl group (CH3 group) to a variety of important metabolic intermediates, including DNA bases and neurotransmitters.
So fundamental is this role of methylation, that the body (essentially, the liver) has numerous pathways of conserving, recycling, and salvaging methionine, so that it can withstand long periods of reduced methionine intake. So intense has been the emphasis on the essentiality of methionine, that methionine deficiencies have been hypothesized to underlie a number of pathologies, including cancer. But nothing could be further from the truth. In fact, typical Westerners consume about 10 times more methionine than is needed to support good health. (We really only need about 300 – 500 mg per day of methionine: more like the daily requirement for a B vitamin than a bulk nutrient!) Over 20 years’ worth of animal research has shown that laboratory animals (rats and mice) live substantially longer and healthier lives if their normal methionine intake is reduced by 80%. Moreover, it is now understood that metabolically, far from salvaging, recycling and regenerating methionine, the body switches gears and gets rid of most of the methionine absorbed from a typical high-protein meal.
What’s that got to do with glycine? The answer is remarkably simple: There is only one metabolic pathway that exists in the human body to get rid of excess methionine. That pathway—via the enzyme glycine-N-methyltransferase (GNMT)—uses up glycine in the process.
In Figure “a†below, we see a typical description of what is called the methionine cycle. It is my own version (I originally published these diagrams at the Annual Meeting of the Federation of American Societies for Experimental Biology [FASEB] in 2011.), but it typically shows how methionine is recycled, in order to conserve it maximally. I say typical, because this is the only metabolic picture generally shown in textbook descriptions of the methionine cycle. And it is accurate insofar as it applies when methionine levels are low, like when there is no methionine coming in from the diet, so methionine needs to be conserved.
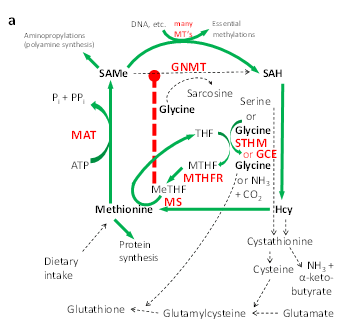

In this metabolic diagram, the green arrows refer to active metabolic pathways, and the dotted black arrows refer to inactive or minimally active pathways; the red, upper case abbreviations stand for enzymes (e.g., GNMT), whereas the names and abbreviations in black are metabolic intermediates (e.g., SAMe). The heavy red dotted line shows a metabolic brake or inhibition (i.e., an “off switchâ€) of GNMT by an intermediate called MeTHF. MeTHF is a form of folic acid which recharges the intermediate amino acid homocysteine (Hcy) by adding a methyl group to it, thus regenerating methionine. The presence of MeTHF is a signal that methionine is being regenerated because it is scarce, and it specifically turns off GNMT so that methionine is not wasted in this time of need. The SAMe that is generated is available for essential processes, such as the methylation of DNA bases, as shown by the green arrow across the top of the diagram.
But what is not generally appreciated by biochemists and nutritionists is that when methionine is abundant, especially when methionine is being absorbed after a high-protein meal, the liver’s metabolic machinery switches gears (like it does in the transition after a high carb meal, when it switches from regenerating glucose to getting rid of glucose), working maximally to get rid of the excess methionine.
This situation is illustrated in Figure “b†below. In this diagram are also shown the two metabolic “on switches†(Solid yellow arrows with star points) for enzymes. Specifically, the high concentration of methionine itself cranks up the activity of MAT, which turns methionine into SAMe at a much higher rate. Most of this SAMe is not needed for essential methylation reactions, such as making DNA bases, but is instead deliberately wasted by GNMT, which has been turned on by the release of the braking action of MeTHF (shown in figure “aâ€). That’s because MeTHF is no longer being made, because SAMe itself shuts off MTHFR, the enzyme that makes MeTHF. Meanwhile, SAMe also turns on the enzyme CBS, which diverts homocysteine (“used†methionine) into the production of downstream sulfur-containing compounds (a pathway called “transulfurationâ€), including cysteine and glutathione, now that the remethylation (regeneration) of methionine has been turned off.

Note especially that in this high-methionine condition, glycine is required both for the action of GNMT (a pathway called “transmethylationâ€) and for the transsulfuration pathway, to make glutathione. In this mode, glycine can be made both from serine and from scratch (i.e., from CO2 and ammonia), through the operation of a glycine-serine cycle. However, the liver cannot keep up with the need for glycine when methionine intake is too high relative to glycine intake (i.e., when we eat lots of muscle meat without the accompanying collagen from the bones and connective tissues), and glycine levels end up being inadequate to properly regulate the immune system. The result: chronic inappropriate and/or excessive inflammation. The antidote: Eat enough glycine to balance the high intake of methionine. 8 grams per day is about right for the typical omnivorous diet.
* * * * *
About the Author: Joel Brind, Ph.D. has been a Professor of Biology and Endocrinology at Baruch College of the City University of New York for 28 years and a medical research biochemist since 1981. Long specializing in steroid biosynthesis and metabolism and endocrine-related cancers, he has specialized in amino acid metabolism in recent years, particularly in relation to glycine and one-carbon metabolism. In 2010 he founded Natural Food Science, LLC to make and market glycine supplement products via http://sweetamine.com

'Diet and Inflammation Part 1-4 – By Joel Brind, Ph.D' has no comments
Be the first to comment this post!